INTRODUCTION
Toxoplasma gondii is a ubiquitous intracellular protozoan parasite that can invade and replicate in all nucleated cells. It is prevalent in humans and animals worldwide, and one-third of the world’s population is reportedly infected with T. gondii [1,2]. Although 80–90% of individuals with primary infection are asymptomatic, the most affected are immunocompromised individuals (HIV/AIDS, cancer patients, organ and tissue transplant recipients, and congenitally infected children), and in these patients, T. gondii can lead to encephalitis, ophthalmopathy, or even death [3–5]. T. gondii infection in domestic animals can cause substantial economic losses due to abortion and neonatal loss in livestock and represents a source of transmission to humans [6–8].
To date, no safe and effective drug is available to eliminate T. gondii tissue cysts [9]. Thus, the development of a T. gondii vaccine is considered to be the most effective way to control toxoplasmosis [10]. Unfortunately, after decades of effort, only one vaccine (Toxovax®) has been licensed for use to prevent abortion in sheep and goats, and no vaccine suitable for use in humans exists [11]. In these circumstances, there is an urgent need to develop new, effective vaccines to reduce the high incidence of toxoplasmosis by preventing and treating this disease in humans and animals [12].
DNA vaccines have become a major focus of research, because they promote the specific expression of an encoded vaccine antigen by host cells and can deliver multivalent vaccines to a host in a single dose. In particular, DNA vaccines can also elicit potent, long-lasting humoral and cell-mediated immunity [13]. T. gondii has evolved unique secretory organelles to help establish infection in its hosts. Among these secretory organelles, the dense granules (GRA), micronemes (MIC), and rhoptries (ROP) are the best characterized, and these organelles play critical roles in adhesion, motility, host invasion, intracellular replication, manipulation of host signaling and innate immune pathways, and formation of the parasitophorous vacuole (PV) [14,15]. The vaccine candidate antigens involved in protective immunity against T. gondii include membrane-associated surface antigens (SAGs) and secreted GRAs, MICs and ROPs [16–24]. However, none of these antigens could completely inhibit tissue cyst formation [25].
Dense granules is an important secretory organelle in the T. gondii cytoplasm, and GRA proteins are potent antigens that trigger strong T and B cell responses upon infection [20,26]. Previous studies have demonstrated that GRA8 is a marker of acute infection, and IgG, IgM or IgA detection with GRA8 by enzyme-linked immunosorbent assay (ELISA) is useful in diagnosing T. gondii infection and discriminating acute from chronic infection [27–29]. However, no study of the protective efficacy of a DNA vaccine expressing T. gondii GRA8 to induce resistance to acute infection has been published. To evaluate the protective immune responses induced by T. gondii GRA8, we constructed a GRA8-expressing eukaryotic plasmid, pDsRed2-GRA8, and evaluated its immunogenicity and protective efficacy after vaccination of BALB/c mice through challenge with the highly virulent T. gondii GFP-RH strain.
MATERIALS AND METHODS
Animals
Specific-pathogen-free (SPF) female BALB/c mice of 6 to 8 weeks old were purchased from Guangdong Medical Laboratory Animal Center, Guangzhou, Guangdong Province, China. All mice were handled in strict accordance with the Good Animal Practice Requirements of the Animal Ethics Procedures and Guidelines of the People’s Republic of China. This study was approved by the Committee on the Ethics of Animal Experiments of Guangdong Medical University (Permit Number: SYXK [Guangdong] 2015-0147).
Toxoplasma gondii strain
The tachyzoites of the T. gondii GFP-RH strain were maintained in human retinal pigment epithelial cells (ARPE-19; ATCC, Rockville, Maryland, USA). Briefly, T. gondii GFP-RH were infected into ARPE-19 cells (parasite/cell ratio, 5: 1) and incubated at 37˚C with 5% CO2 for 2 to 3 days. Following host cell rupture, lysed tachyzoites and host cellular debris were centrifuged at 900×g for 10 min using Percoll (Sigma Chemical Co., St. Louis, Missouri, USA) to pellet the parasites. The final pellet was suspended in cold phosphate buffered saline (PBS), and the suspension was passed through a 5.0-μm-poresize filter (Millipore, Bedford, Massachusetts, USA). Purified tachyzoites were used in all experiments.
Preparation of soluble tachyzoite antigen
The soluble tachyzoite antigens (STAg) were prepared from T. gondii tachyzoites as described previously [26]. Briefly, purified tachyzoites were centrifuged at 5,000×g for 3 min and disrupted by 3 cycles of freezing at −20˚C and thawing at 4˚C. Finally, the lysate was sonicated on ice at 60 W/s and centrifuged for 40 min at 100,000×g. The supernatants containing STAg were pooled and sterile filtered (Millipore), and the protein concentration was determined via the Bradford method using bovine serum albumin (BSA) as the standard. STAg was stored in aliquots at −80˚C until used.
Construction and preparation of DNA vaccine plasmid
The total RNA of the RH strain tachyzoites was prepared by using TRIzol reagent (Invitrogen Life Technologies, Carlsbad, California, USA) according to the manufacturer’s instructions. The coding sequence of GRA8 was obtained by RT-PCR amplification from total RNA with specific primers (forward primer: 5′-GGC CTC GAG ATG GCT TTA CCA TTG CG-3′ and reverse primer: 5′-CCG GAA TTC GCG TCG TTA CGG TGA ATC-3′, containing the XhoI and EcoRI restriction sites, respectively, which are underlined) and then inserted into the pGEM-T Easy vector (Promega Corporation, Madison, Wisconsin, USA). The GRA8 fragments were cleaved from pGEM-GRA8 by using the restriction endonucleases XhoI and EcoRI and then were subcloned into the corresponding sites of the pDsRed2-N1 vector (Invitrogen Life Technologies) to construct pDsRed2-N1-GRA8 (pDsRed2-GRA8). The recombinant plasmid was propagated in Escherichia coli DH5α and confirmed by restriction analysis and PCR sequencing (Sangon Biotech, Shanghai, China).
Large-scale plasmid DNA was prepared using the Endotox in-Free Mega kit according to the manufacturer’s instructions (Qiagen, Hilden, Germany); concentrations were determined by spectrophotometer, and the DNA was diluted with sterile endotoxin-free PBS to a final concentration of 1 mg/ml and stored at −20˚C until use.
Expression of pDsRed2-GRA8 in vitro
The recombinant plasmids pDsRed2-GRA8 and pDsRed2-N1 were separately transfected into ARPE-19 cells grown on glass coverslips in 12-well plates by using Lipofectamine LTX and Plus reagents (Invitrogen Life Technologies) according to the manufacturer’s instructions. Forty-eight hours after transfection, cells were fixed with 4% paraformaldehyde and per-meabilized with 0.1% Triton X-100 in PBS (PBST) for 10 min. The coverslips were mounted on microscope slides using a mounting medium containing 4′,6-diamidino-2-phenylindole (DAPI) (Vector Laboratories, Burlingame, California, USA), and the cells were imaged using fluorescence microscopy.
DNA immunization and experimental design
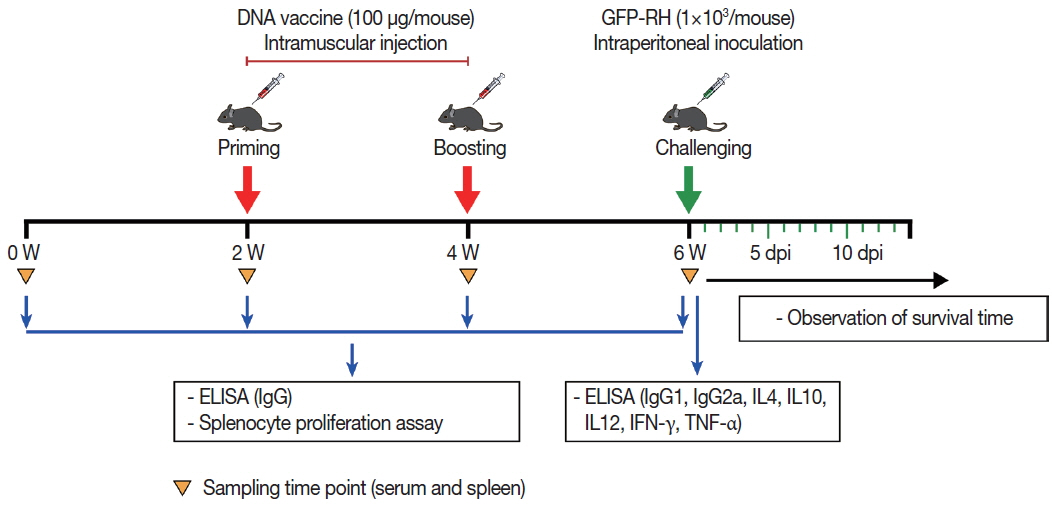
A total of 80 female BALB/c mice were randomly divided into 4 groups (20 mice per group). For the experimental groups, mice were inoculated with an injection of 50 μg pDsRed2-GRA8 into the tibialis anterior muscles of both hind legs (100 μg/per mouse) and given booster immunizations 2 and 4 weeks later. Mice injected with empty pDsRed2-N1vector or sterile endotoxin-free PBS served as negative control groups, and the blank control group received nothing.
Serum samples collected from the tail vein from 3 mice in each group at weeks 0, 2, 4, and 6 were centrifuged at 3,000×g for 10 min and stored at −20˚C for later ELISA detection.
Spleens from 3 mice in each group at week 6 were collected under aseptic conditions and used for in vitro splenocyte proliferation and cytokine assays (IL-4, IL-10, IL-12, IFN-γ, and TNF-α).
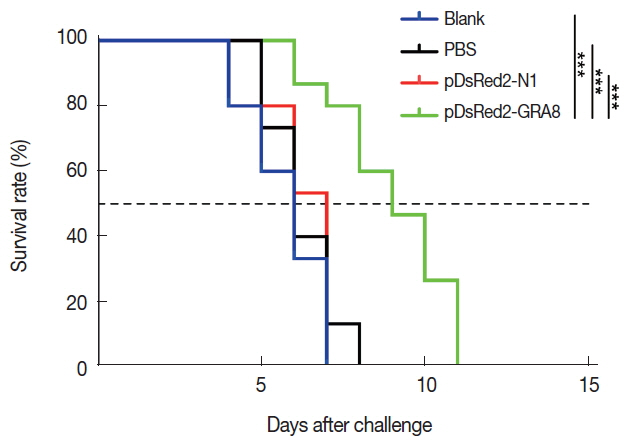
Two weeks after the last immunization, 15 mice per group were challenged intraperitoneally with 1×103 tachyzoites of the highly virulent T. gondii GFP-RH strain, and the survival time of the mice was monitored daily (Fig. 1).
Determination of serum antibodies
T. gondii-specific serum IgG, IgG1, and IgG2a antibody levels were determined by ELISA, as described previously, with some modifications [26]. Briefly, the 96-well plates were coated with 100 μl of STAg (10 μg/ml) in 50 mM carbonate buffer (pH 9.6) at 4˚C overnight. Then, the plates were blocked with PBS containing 1% BSA for 2 hr at room temperature and incubated with the diluted sera. Subsequently, the plates were incubated with horseradish peroxidase (HRP)-conjugated anti-mouse IgG, IgG1, or IgG2a, and the bound antibody was reacted with 200 μl of substrate solution in the dark for 30 min. Finally, the reaction was stopped with 3N HCl, and the absorbance was measured with a spectrophotometer at 490 nm. All samples were evaluated in triplicate.
Splenocyte proliferation assay
Spleens from immunized mice were collected under aseptic conditions in RPMI 1640 medium (Invitrogen Life Technologies). Erythrocytes were removed by erythrocyte lysis buffer (Sigma), and the remaining cells were washed and suspended in RPMI 1640 medium supplemented with 10% FBS, 1% antibiotic-antimycotic (Gibco-BRL, Life Technologies Ltd, Paisley, UK), HEPES (10 mM), L-glutamine (2 mM), sodium pyruvate (1 mM), and β-mercaptoethanol (50 μM). Splenocytes were stimulated with STAg (10 μg/ml) or medium alone as positive and negative controls, respectively, and then incubated at 37˚C in 5% CO2. Splenocyte proliferation was measured using a chemiluminescent bromodeoxyuridine (BrdU) ELISA kit according to the manufacturer’s instructions (Roche Molecular Biochemicals, Indianapolis, Indiana, USA).
Cytokine assays
Splenocytes from each of 3 mice per group were harvested as described above. Single-cell suspensions of splenocytes were stimulated with STAg or medium alone (negative control) in flat-bottomed 96-well microtiter plates as described for the lymphocyte proliferation assay. Culture supernatants were harvested at 24 hr for the determination of IL-4, 72 hr for IL-10, 96 hr for IL-12 (p70) and IFN-γ, 4 and 8 hr for TNF-α, using commercial ELISA kits according to the manufacturer’s instructions (R&D Systems, Minneapolis, Minnesota, USA). Cytokine concentrations were determined by reference to standard curves constructed with known amounts of mouse recombinant IL-4, IL-10, IL-12 (p70), IFN-γ or TNF-α. The sensitivity limits for the assays were 2 pg/ml for IL-4, 1.97 pg/ml for IL-10, 5.0 pg/ml for IL-12 (p70), 2 pg/ml for IFN-γ, and 1.88 pg/ml for TNF-α.
Statistical analysis
Results are presented as the means±standard deviation (SD) of 3 independent experiments. The statistical evaluation of the differences in survival rates was checked by the Kaplan-Meier test, and then a log-rank test was performed. Statistical differences in parasite burdens, antibody titers, in vitro splenocyte proliferation, and cytokine levels were determined by one-way ANOVA with Dunnett’s multiple comparisons test. P-values lower than 0.05 were considered statistically significant.
RESULTS
Expressed recombinant T. gondii GRA8
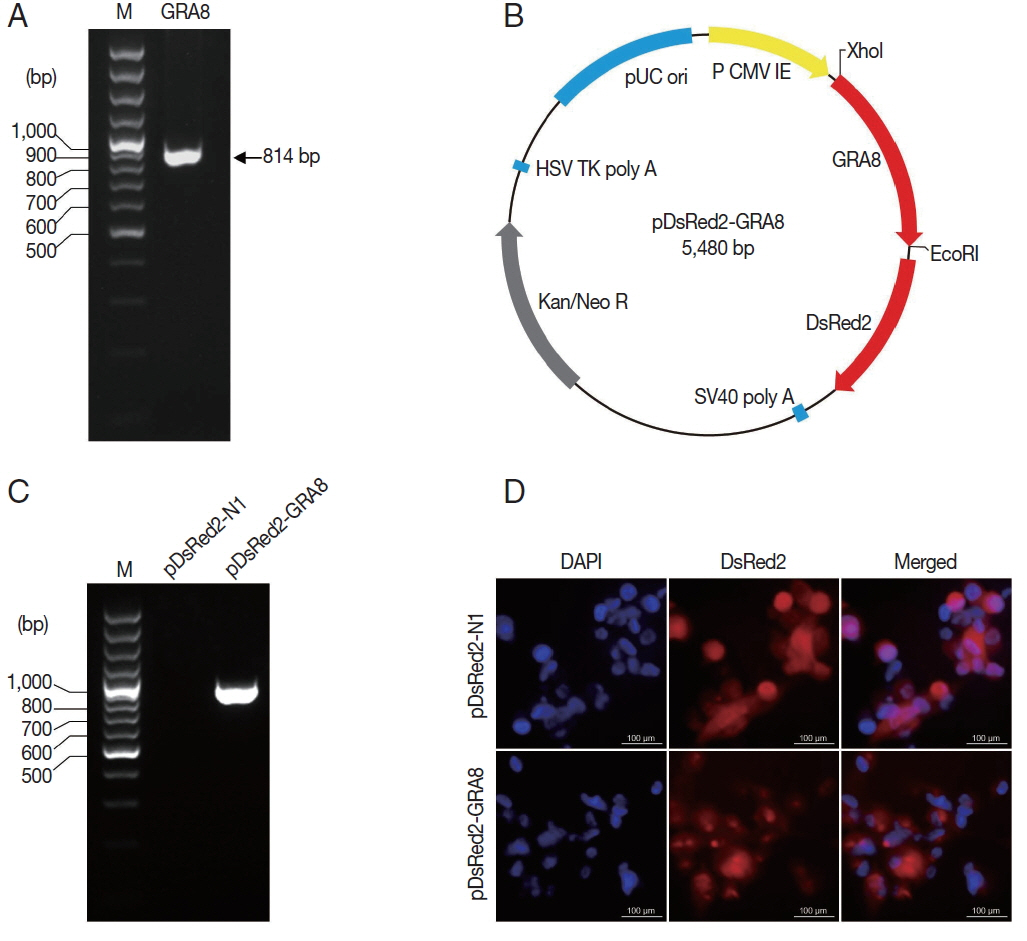
The T. gondii GRA8 gene was amplified by PCR (814 bp) from the cDNA of T. gondii strain GFP-RH (Fig. 2A). Subsequently, the PCR products were digested with the appropriate restriction enzymes and subcloned into the XhoI and EcoRI sites of the pDsRed2-N1 expression vector to construct a recombinant plasmid, which was named pDsRed2-GRA8 (Fig. 2B). To identify the expression of the recombinant plasmid in vitro, the presence of GRA8 mRNA in transfected ARPE-19 cells was analyzed using RT-PCR. As shown in Fig. 2C, no expression of GRA8 was detected in pDsRed2-N1 transfected cells, whereas the expression of GRA8 was detected in pDsRed2-GRA8 transfected cells. In addition, we also confirmed the expression of pDsRed2-GRA8 by immunofluorescence assay. DsRed2 fluorescence was observed in the cytoplasm of ARPE-19 cells transfected with the pDsRed2-N1 vector or pDsRed2-GRA8 (Fig. 2D). These results indicated that the T. gondii GRA8 was expressed successfully in ARPE-19 cells.
Humoral immune responses induced by DNA immunization
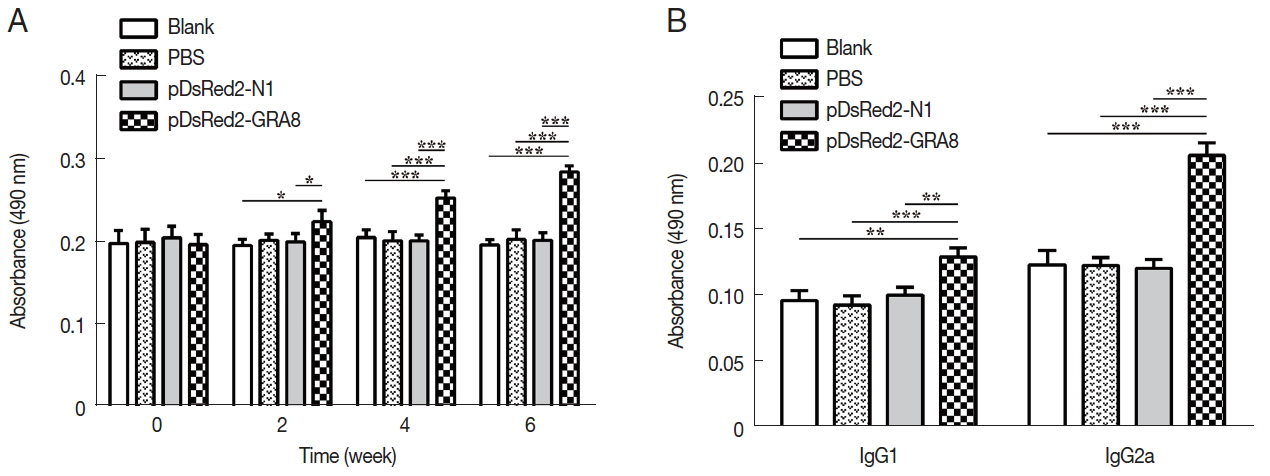
Serum samples from immunized and control mice were used to evaluate the levels of specific antibodies against T. gondii. As shown in Fig. 3A, immunization induced a significant IgG response in all immunized mice from 2 weeks after immunization, with higher antibody levels in pDsRed2-GRA8 immunized mice than in the blank, PBS, or pDsRed2-N1 control groups (2 weeks: F(3,8)=5.082, P=0.0294; 4 weeks: F(3,8)=21.21, P=0.0004; 6 weeks: F(3,8)=66.82, P <0.0001). In contrast, the levels of IgG antibodies in the mice of the 3 control groups did not statistically increase with successive immunizations.
To characterize whether a Th1 and/or Th2 response was induced in the immunized mice, the distribution of subclasses of IgG (IgG1 and IgG2a) was analyzed individually in the sera of mice from all groups at 2 weeks after the last immunization. As depicted in Fig. 3B, the levels of IgG1 and IgG2a in the immunized groups were significantly higher than those in the 3 control groups (IgG1: F(3,8)=17.05, P =0.0008; IgG2a: F(3,8)= 72.47, P<0.0001). The ratios of IgG2a/IgG1 in mice immunized with pDsRed2-GRA8 (1.60±0.11) were significantly different from those in the control groups (blank, 1.29±0.19; PBS, 1.28±0.15; pDsRed2-N1, 1.19±0.81) (F(3,8)=4.908, P=0.032). These results suggested that a specific humoral response with a mixed Th1/Th2 profile was elicited by these DNA immunizations, with a predominant Th1-type immune response in mice immunized with pDsRed2-GRA8.
Splenocyte proliferation
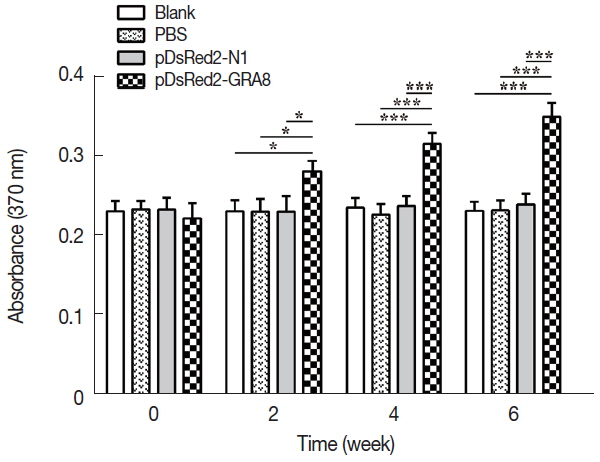
To assess proliferative immune responses to STAg, the splenocytes from the immunized and control groups (3 mice per group) were analyzed by a BrdU ELISA kit. As shown in Fig. 4, in vitro splenocyte proliferation in mice immunized with pDsRed2-GRA8 was significantly higher than that in the 3 control groups (2 weeks: F(3,8)=7.595, P =0.01; 4 weeks: F(3,8)=32.32, P <0.0001; 6 weeks: F(3,8)=51.46, P <0.0001) and gradually increased over time starting 2 weeks after immunization. In contrast, the in vitro splenocyte proliferation among the mice in the 3 control groups did not show significant differences.
Cytokine production
To determine whether pDsRed2-GRA8 immunization augments the Th1 or Th2 cytokine response, STAg-treated culture supernatants of splenocytes were obtained from immunized mice 2 weeks after the final immunization, and the levels of IL-4, IL-10, IL-12 (p70), IFN-γ and TNF-α were quantified by ELISA. As shown in Table 1, the levels of IL-10, IL-12 (p70), IFN-γ, and TNF-α in spleen cell cultures from mice immunized with pDsRed2-GRA8 were significantly increased compared to those in the 3 control groups (IL-10: F(3,8)=38.72, P <0.0001; IL-12: F(3,8)=224.4, P <0.0001; IFN-γ: F(3,8)=219.8, P <0.0001; TNF-α: F(3,8)=88.22, P <0.0001). However, the level of IL-4 in spleen cell cultures from the pDsRed2-GRA8 immunized group was not significantly higher than those in the 3 control groups.
Protection of immunized mice
To evaluate the protective immunity induced by immunization with pDsRed2-GRA8, 15 mice from each group were challenged intraperitoneally with 1×103 tachyzoites of the virulent T. gondii GFP-RH strain at 2 weeks after the last immunization. The survival times of the mice were observed. The average survival time of the mice immunized with pDsRed2-GRA8 (9.00 ± 1.77 days) was significantly longer than that of the 3 control groups (blank, 5.73±1.16 days, χ2=20.43, df=1, P <0.0001; PBS, 6.27±1.03 days, χ2=17.34, df=1, P <0.0001; pDsRed2-N1, 6.33±0.82 days, χ2=17.31, df=1, P <0.0001). However, the difference in the average survival time of mice among the 3 control groups was not statistically significant, and all died within 8 days after challenge (Fig. 5).
DISCUSSION
In recent years, owing to the easy production and low cost of DNA vaccines and the fact that DNA vaccines can elicit long-lasting humoral and cellular immune responses, the development of effective DNA vaccines has emerged as an active research field in the fight against toxoplasmosis [22,30,31]. Numerous studies have evaluated the protective efficacy of different antigens, such as SAG4, GRA15, MIC13, ROP18, NTPase II, TrxLp, and ENO2 [18,24,32–35]. Most of them showed partial protection against toxoplasmosis. Herein, we evaluated the protective immunity induced by DNA vaccination with a plasmid encoding GRA8 proteins against acute toxoplasmosis in a mouse model. Our results demonstrated that immunization with pDsRed2-GRA8 can trigger different levels of humoral and cellular responses, with a slightly longer survival time than the blank, PBS or pDsRed2-N1 controls, suggesting that T. gondii GRA8 DNA immunization induces a partial protective effect against acute toxoplasmosis.
When T. gondii invasion occurs for the first time in vivo, the parasite can be captured and processed by antigen-presenting cells and then presented to T lymphocytes, which further build adaptive immunity. When T. gondii invades subsequently, special anti-T. gondii IgG antibodies adhere to the surfaces of the parasites and limit their spread by preventing attachment to host cell receptors, resulting in their elimination by macrophages [36]. Therefore, humoral response, by promoting macrophages to kill intracellular parasites, has been considered to be of great importance in immunity against T. gondii infection. In the present study, we evaluated humoral response intensity on the basis of specific anti-T. gondii IgG levels. The immunization of mice with pDsRed2-GRA8 could induce significantly higher levels of IgG antibodies than those in the controls (P <0.05), which would contribute to strong protective efficacy against T. gondii infection. These results were in agreement with those of previous studies that vaccinated mice with plasmids encoding GRA7, GRA17, GRA23, ROM5, and ROP38 [10,26,37,38].
High levels of IgG1 and IgG2a were also detected in the serum of mice in the pDsRed2-GRA8 immunized group compared to those in the control groups. This result was consistent with those of previous studies, showing that DNA immunization could elicit a mixed Th1/Th2 immune response, with a more significant IgG2a response [10,39–41]. The higher ratio of IgG2a/IgG1 in mice immunized with pDsRed2-GRA8 shows that a predominantly Th1-type immune response was elicited.
A Th1-type immune response associated with high levels of IFN-γ is essential for protection against T. gondii infection [42]. By restimulation of splenocytes in vitro with STAg, significant increases in IFN-γ, IL-12 (p70) and TNF-α production were induced after DNA immunization with pDsRed2-GRA8. These results were similar to previous data showing that immunization with a plasmid encoding the GRA7 gene enhanced the production of IFN-γ in vitro and GRA7-specific IgG2a antibody [26]. These results suggested that immunization with a pDsRed2-GRA8 DNA vaccine could elicit the Th1-type immune response that contributes to effective protection against acute T. gondii infection. In addition, the Th2-type cytokines (IL-4 and IL-10) have been shown to play important roles during the early phase of acute T. gondii infection [43]. In our study, increased levels of IL-10 were observed in only the mice immunized with pDsRed2-GRA8. As a regulatory cytokine, IL-10 is important for protection against the potential immunopathological mechanism caused by a vigorous Th1 response characterized by high levels of IFN-γ production [44]. Therefore, the high level of IL-10 contributed to the longer survival time of mice immunized with pDsRed2-GRA8. However, the levels of IL-4 in all immunized groups were similar to those in all control groups. These results may further explain the inability of these DNA vaccines to completely protect mice against acute T. gondii infection.
Cellular immunity plays an important role in the control of T. gondii infection. In this study, a significantly higher level of splenocyte proliferative response was induced by DNA immunization with pDsRed2-GRA8. This result was similar to those of previous studies reporting that DNA vaccination with GRA7 and GRA16 could increase splenocyte proliferation compared with that in controls [25,26]. These results suggest that a cellular immune response was elicited in the immunized mice. Immunization with pDsRed2-GRA8 induced partial immune protection against acute toxoplasmosis, and all immunized mice had prolonged survival times compared to those of all controls. These results indicate that pDsRed2-GRA8 represents a potential candidate for the further development of effective vaccines against acute toxoplasmosis. Nevertheless, further studies are needed to assess the induction of stronger immune protection by multiantigenic DNA vaccines combining GRA8 with other proteins to combat acute and chronic toxoplasmosis. The present study showed that DNA immunization with pDsRed2-GRA8 induced humoral and cellular immune responses, and all immunized mice showed greater Th1-type immune responses and longer survival times than those of control mice. However, the changes of immune factors by pDsRed2-GRA8 immunization just had a little effect on the survival of acute toxoplasmosis mice. These results indicated that T. gondii GRA8 DNA immunization induces a partial protective effect against acute toxoplasmosis.